
Tato známá zelenina opraví vše, co ve vašem těle nefunguje
Existuje jedna extrémně zdravá zelenina s mocnými medicínskými vlastnostmi, která dokáže přinést úlevu a vyléčení z více nemocí a zdravotních problémů. Za své účinky vděčí převážně vysokému obsahu látek známých pod názvem anthokyaniny, které jí dodávají typické červené zbarvení. Kromě jiných věcí mají i nesmírně silné...

Promývání úst olejem: Věda právě potvrdila to, co Indové věděli už dávno
Tvrdí se, že tato metoda bělí zuby, udělá dech z úst svěžejší a celkově vede k masivnímu zlepšení orální hygieny....

Věda potvrdila, že kurkuma je účinnější než 15 druhů léků
Kurkuma patří v současnosti mezi nejvíce zkoumané rostliny. Medicínské účinky v ní obsažených látek (primárně...

Doktor mi nařídil spát na zádech a bez polštáře, tento trik se učím dělat každou noc
Během spánku tělo obnovuje životně důležité zdroje. K tomu by se zdálo, že stačí spát 6–8 hodin denně. Ne každý dříč...
Populární příspěvky






Blog

Zázračný propolis. Pomůže při zánětech žaludku nebo žaludečních vředech
Včelí tmel je zvláštní lepkavá substance vytvářená včelami za účelem řešení nejrůznějších každodenních úkolů. Pokud například v úlu vznikne díra, nebudou včely trpět zimou, protože ji jednoduše zaplní propolisem....

Až vyzkoušíte tento lék, už nikdy nebudete cítit bolesti kloubů a kostí!
Mnoho vědců a zdravotnických výzkumníků zjistilo, že hlavním důvodem proč je bolest zad, nohou a bolesti kloubů je nesprávné držení těla. Takže, pokud jste setkali s takovými bolestmi v kloubech, nohách a krku pak...

Lepší než léky od lékaře: Mocný čaj na oteklé nohy
Petržel vypadá dobře jako okrasa jídel. Mnozí však už zapomněli, že tato nádherná rostlinka může být využita i na spoustu jiných účelů. Konzumace dostatečného množství petržele se vždy spojovala se zdravím. Věří...

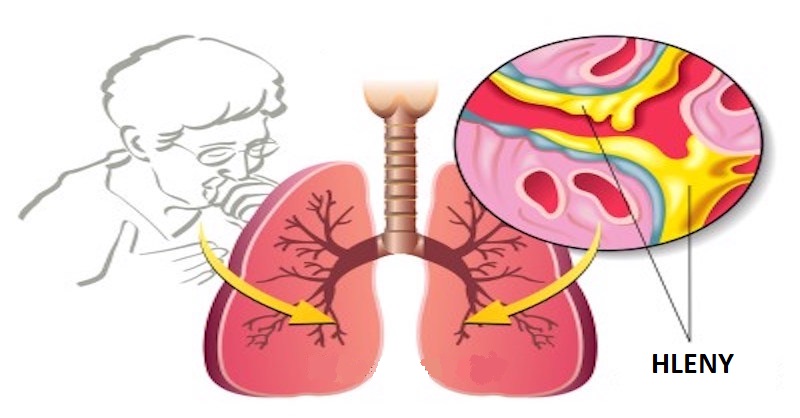
7 mocných domácích léčebných terapií na zánět plic (pneumonii) – zmizí za 3 dny
Pneumonie je respirační onemocnění, které je charakterizované zánětem plic, které způsobují bakterie, viry nebo chemická dráždidla. Tělo obvykle brání těmto organismům infikovat vaše plíce. Nicméně někdy to váš...

Citron a jedlá soda – kombinace, která zachraňuje životy!
Předpokládá se, že kombinace citronu a jedlé sody má 1000krát větší účinek než chemoterapie! Proč je tato skutečnost tak dlouho utajována? Četné studie prokázaly, že citron má proti rakovinné...

S tímto nápojem dokonale posílíte imunitu a detoxikujete játra
Jistě si pamatujete, že jste v dětství před spaním pili sklenici horkého mléka, což vám pomohlo rychle usnout.Prozradíme vám recept na podobný nápoj, který před spaním uklidní vaše tělo i mysl a poskytne vám celou řadu...
Japonský způsob, jak odstranit všechny škodlivé toxiny z těla. Přirozeně!
Kdysi byly detoxikační podložky v Japonsku používány pro odstranění škodlivých toxinů z těla. Dnes už lidé nemají tendenci je používat, ale musíme říci, že jsou neuvěřitelně efektivní. Jediné, co musíte udělat, je...
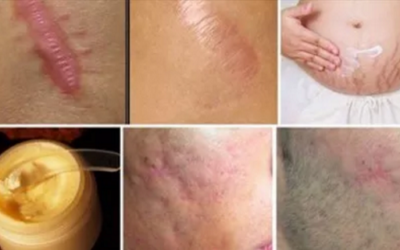
Zbavte se do měsíce jizev na jakékoliv části vašeho těla
Můžeme vám položit jednoduchou otázku? Víte co je to vlastně jizva? Ano, jizva je značka, kterou zanechá úraz nebo poranění na vaší pokožce poté, co dojde k její zahojení. Všichni víme, že jizvy jsou poměrně...

Léčba – pitím čisté vody. Přemrzlá voda – jednoduchá detoxikace těla
MUDr. F. Batmanghelidj - M. Adams: exklusivní neortodoxní rozhovor reportéra M. Adamse s neortodoxním Dr. F. Batmanghelidjem, autorem knihy Water for Health, For Healing, For Life (Voda pro zdraví, léčbu,...